A concentração micelar crítica refere-se ao teor de um surfactante mínimo acima do qual esse pode reorganizar-se em micelas ou lipossomos. É bastante utilizado na caracterização de tais compostos, como biosurfactantes na indústria, e pode ser medido por diversas técnicas, incluindo tensão superficial, fluorescência de polarização, turbidimetria e absorção molecular (fotometria).
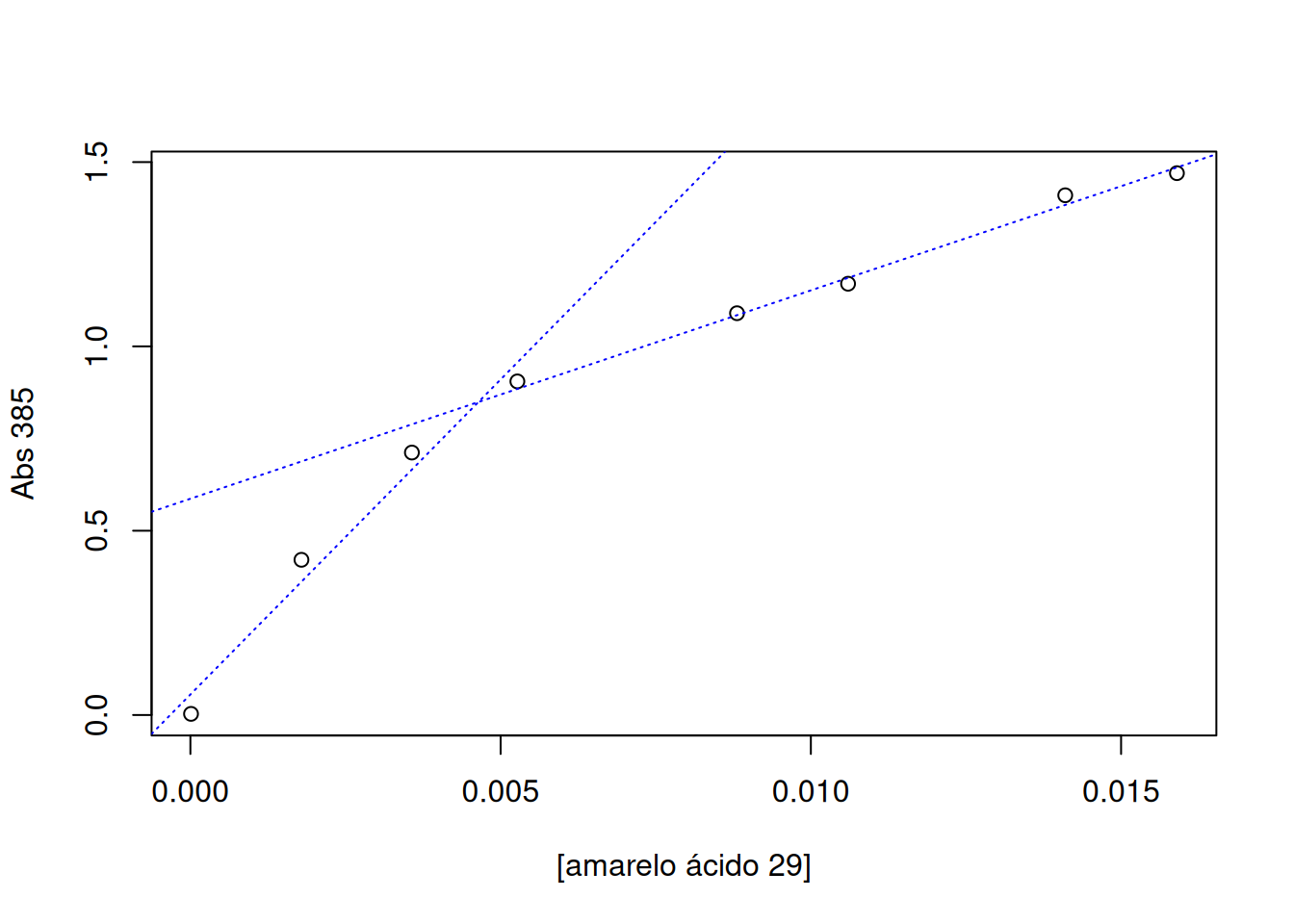
De modo geral a determinação de cmc é obtida pelo valor da concentração do surfactante no ponto de cruzamento de duas retas obtidas por ajuste linear dos dados em baixo e alto teor do analito, como segue:
O ponto de intersecção pode ser obtido manualmente, pelo comando locator já referido, ou de forma automática. Nesse caso, o exemplo do R que ilustra esse cálculo baseia-se em resultados de espectrofometria obtidos para um corante, o amarelo ácido 29 (Duff e Giles 1972).
# Determinação de concentração micelar crítica (CMC)conc <-c(1e-5, 0.00179, 0.00357, 0.00527, 0.00881, 0.0106, 0.0141, 0.0159) # teor do corante (mol/L)A385 <-c(0.003, 0.421, 0.712, 0.905, 1.09, 1.17, 1.41, 1.47) # absorbância em 385 nm# Gráficoplot(A385 ~ conc, xlab ="[amarelo ácido 29]", ylab ="Abs 385")# Ajuste linear para 2 conjuntos de dados# 1o. conjuntolinCmc1 <-lm(A385 ~ conc, subset = (conc <0.007& conc >0)) # 1o. ajuste linear com limitesabline(linCmc1, col ="blue", lty ="dotted") # linha de regressão# 2o. conjunto:linCmc2 <-lm(A385 ~ conc, subset = (conc <0.02& conc >0.007)) # 2o. ajuste linear com limitesabline(linCmc2, col ="blue", lty ="dotted") # linha de regressão
# Cálculo de CMC por intersecção das automática das duas retas:cmc_auto <-abs((coef(linCmc2)[1] -coef(linCmc1)[1]) / (coef(linCmc1)[2] -coef(linCmc2)[2]))as.numeric(cmc_auto) # fornece o cmc em mol/L
[1] 0.004642773
O valor encontrado pelos autores foi de 0,004 mol/L.
Transporte em membranas e Teoria Quimiosmótica
O equilíbrio de transporte de solutos por membranas envolve um formalismo que abrange o potencial eletroquímico dos solutos envolvidos, suas concentrações (ou atividade), cargas, potenciais elétricos e volumes parciais em unidade molal. À despeito dessa complexidade, contudo, podemos ilustrar o transporte de íons H\(^{+}\) de modo simplificado, seguindo-se a equação abaixo:
\[
\Delta G_{transp}=2.303(RT*log\frac{H^+_{in}}{H^+_{out}}) + z * F * \Delta \phi
\]
Onde F representa a constante de Faraday, 96485 J\(^{-1}\)\(V^{-1}\)\(mol^{-1}\) (também representado como 1 mol de elétrons), e z a carga do íon equanto que \(\Delta\)\(\phi\) representa a variação de potencial elétrico, e \(\Delta\)pH a variação do valor de pH, ambos obtidos por medições entre o lado interno (matriz mitocondrial) e externo (espaço intermembranas). H\(^{+}\)\(_{in}\) e H\(^{+}\)\(_{out}\) representam o teor de prótons do lado interno e externo da membrana, respectivamente.
Agora, considerando a carga unitária de H\(^{+}\) e a definição para pH (-log H\(^{+}\)),
Tangente ao transporte de solutos e íons por membranas celulares, é possível prever-se, por exemplo, o teor de ATP formado durante a fosforilação oxidativa que envolve o retorno de íons H\(^{+}\) do espaço intermembranas à matriz mitocondrial. Ilustrando-se, considerando um valor de \(\Delta\)\(\phi\) de 70 mV e um \(\Delta\)pH de 1,4 para as medidas entre matriz e espaço intermembranas mitocondriais, prevê-se a obtenção de ATP pelas relações que seguem, considerando a energia de 31 kJ/mol de ATP :
# Teor previsto de ATP durante fosforilação oxidativaR <-8.341# J/molT <-298# KF <-96485# constante de FaradayDphi <-70e-3# variação de potencial elétrico in/out membranasDpH <--1.4# variação de pH in/out membranasDG_transp <- F * Dphi -2.303* R * T * DpH # equação de transporteDG_transp_4 <-4* DG_transp # 4 mol de H+# Considerando cada mol de ATP para 31 kJ/mol...DG_transp_4 /31e3
[1] 1.905559
Percebe-se, portanto, a produção de 2 mols de ATP nas condições explicitadas.
Proteínas de transporte em membranas
Enquanto prótons como H\(^{+}\) são transportados em função de seu gradiente de concentração entre o lado interno e externo de membranas, outros compostos e solutos dependem de uma proteína transportadora, como glicose e ácido cítrico. Nesse caso, o transporte não é passivo, mas de difusão facilitada, e seu comportamento cinético pela membrana obedece o formalismo de Michaelis-Menten como segue.
Onde, analogamente, V\(_{max}\) representa a velocidade máxima (ou limite) de transporte do substrato, S\(_{out}\) o teor do substrato transportado, e K\(_{transp}\) a constante de dissociação do complexo proteína-substrato (ou concentração do substrato a meia saturação do transportador).
Duff, DG, e CH Giles. 1972. «Spectrophotometric determination of the critical micelle concentration of surfactants». Journal of Colloid and Interface Science 41 (3): 407–14.