8 Cinética Enzimática
8.1 Introdução
9 Introdução
Enzimas ou moléculas com comportamento enzimático (RNA catalítico, por ex) podem ser observadas quanto à sua estrutura, mecanismo de ação, e comportamento cinético. Uma equação que define o comportamento de uma enzima E sobre seu substrato S é:
Nessa equação, P representa o produto da reação, ES o complexo ativado no estado de transição, e k1, k2 e k3 as constantes de velocidade da reação.
9.1 Detalhes
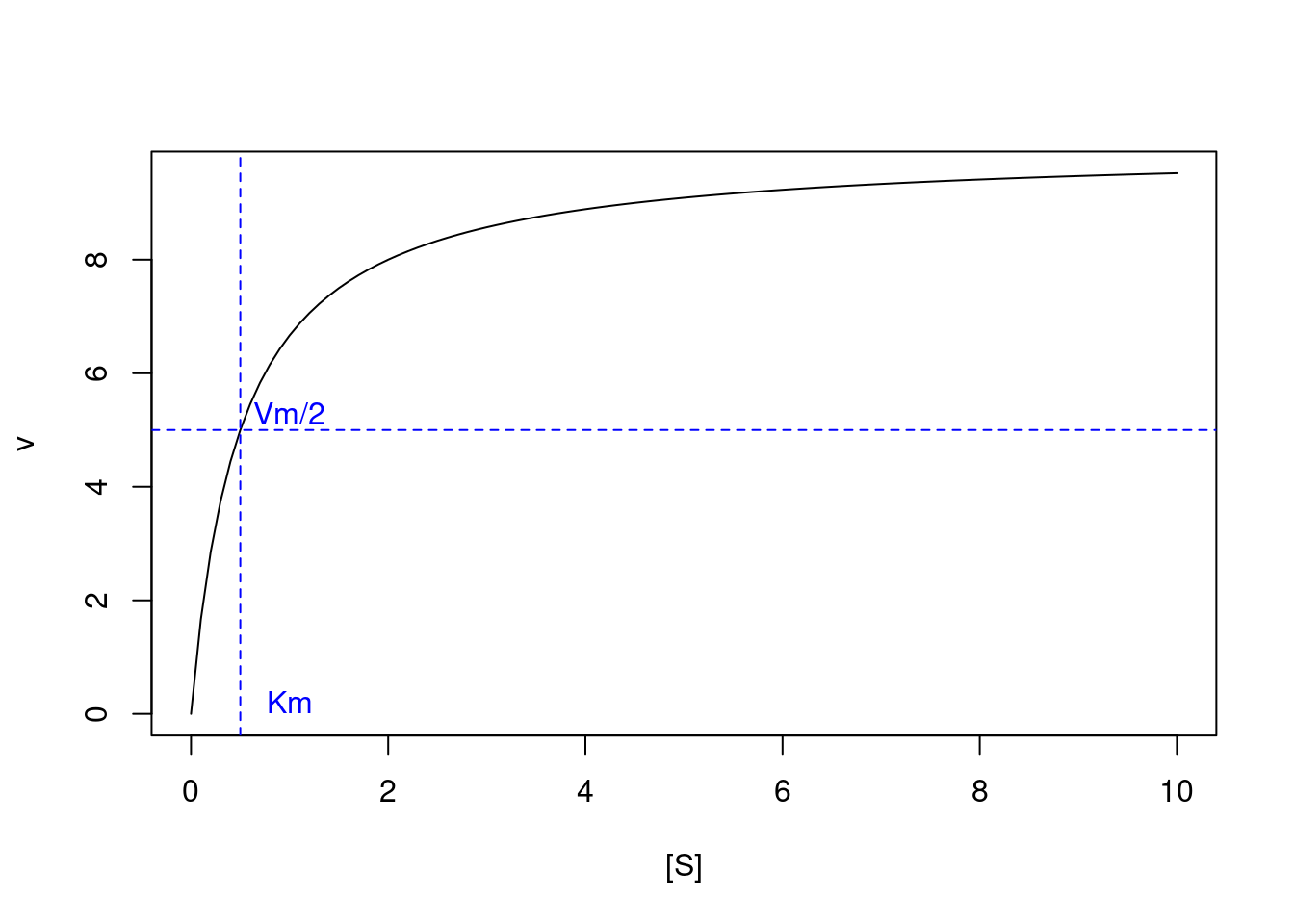
Aplicando-se alguns formalismos, tais como a aproximação de Briggs-Haldane para o estado estacionário, e o tratamento de Henri-Michaelis-Menten, pode-se obter uma relação matemática que define a curva de velocidade da reação enzimática com o aumento do substrato, como segue:
Nessa equação, Km representa a constante de Michaelis-Menten, e Vm a velocidade limite da reação (ou velocidade máxima, como trazem os livros clássicos na área).
A constante Km, por sua vez, pode ser definida a partir das constantes de velocidade da equação @ref(eq:diagMM) como:
Por essa relação, o valor de Km é representado pelo teor de substrato que confere metade do valor de Vm para a reação. Observe que pela equação o valor de Km no denominador influencia inversamente a velocidade v da reação; ou seja, quanto maior o valor de Km, menor a taxa da reação enzimática.
Ainda que a obtenção dos parâmetros cinéticos de uma enzima seja bastante evidente pelo gráfico acima, na prática a assíntota da curva, ou seja, a região de teor de substrato em que ela atinge um ponto máximo, é mais difícil de ser visualizada.
Isso decorre de várias situações, tais como a quantidade insuficiente de substrato para se atingir um máximo, ou efeitos de viscosidade em alta concentração de substrato (reduzindo, portanto, a velocidade da reação), ou mesmo a ação desse como inibidor da enzima, quando em excesso, dentre várias outras situações.
Dessa forma, busca-se obter os parâmetros cinéticos Km e Vm por linearizações, ou seja, transformações matemáticas que visem obter uma reta, ao invés de uma curva.
Uma reta é bem mais simples de ser compreendida que uma curva. Curvas podem ser hiperbólicas (como a de *Michaelis-Menten), exponeciais, de potência, sigmoidais, e outros tantos tipos. Cada qual é representado por uma equação específica.
Já retas são bem mais simples, já que possuem apenas um intercepto (a) em cada eixo, e uma inclinação (b), definidas genericamente por:
Dessa forma a conversão de uma curva de Michaelis-Mentem em reta facilita bastante o cálculo dos parâmetros cinéticos, linearizando a curva homônima.
9.2 Linearizações
Diversas são as linearizações encontradas na literatura para a equação de Michaelis-Mentem. As três transformações mais empregadas são exemplificadas abaixo, juntamente com a curva direta de Michaelis-Mentem.
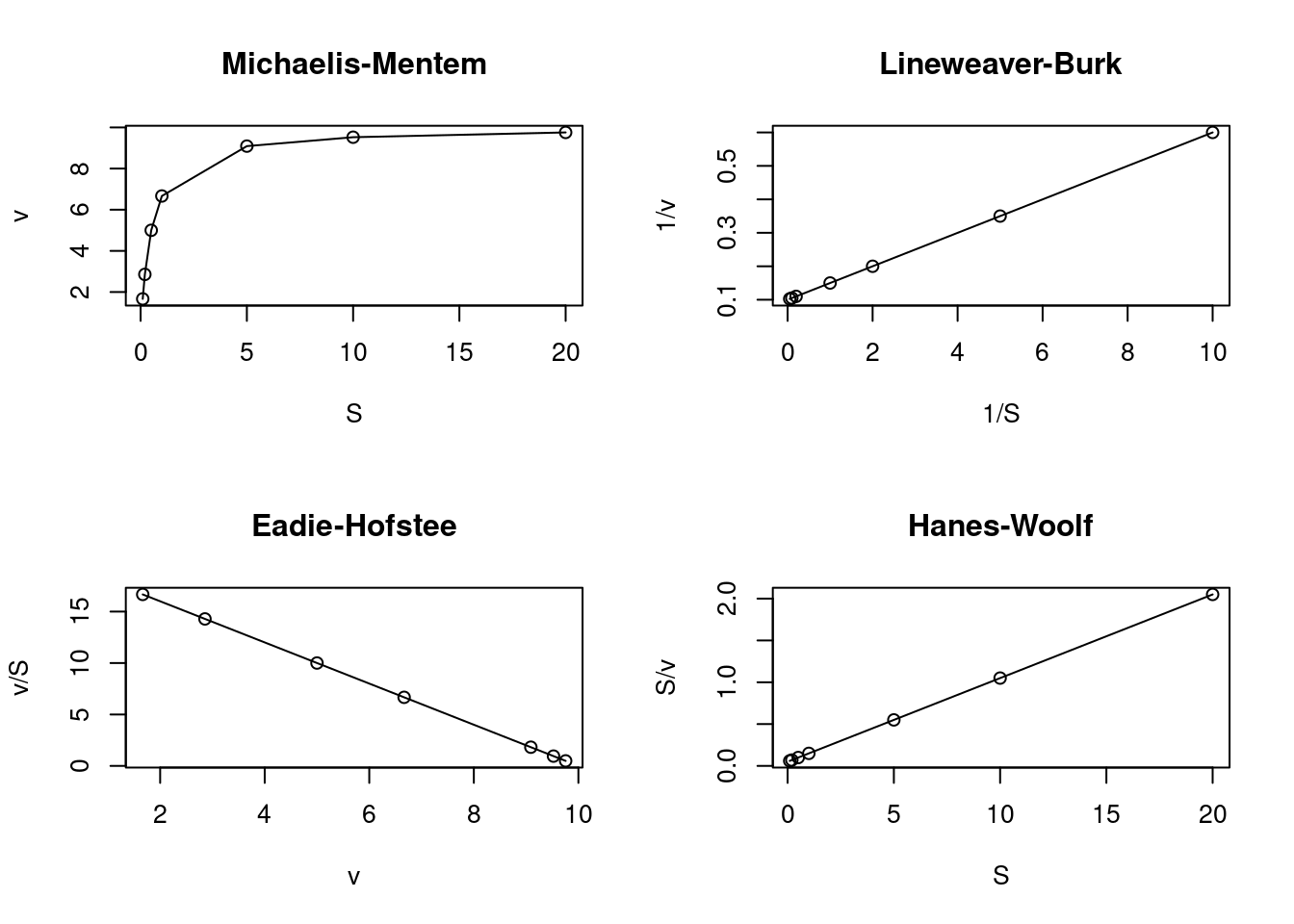
Dentre essas transformações lineares para cinética enzimática, a mais frequentemente encontrada nos textos afins é a de Lineweaver-Burk (também conhecido como gráfico duplo-recíproco), e cuja equação é dada abaixo:
9.2.1 Enzimas alostéricas
O alosterismo constitui um dos principais recursos da metabolismo para a regulação dos níveis de compostos celulares. De etiologia grega (allos = outro, stereos = estrutura), uma enzima alostérica é aquela que altera seu perfil catalítico em função de transições conformacionais mediadas por moléculas que interagem fora de seu sítio ativo, sejam elas substrato, coenzimas, ou outros compostos.
O efeito resultante é uma modulação da atividade enzimática, quer ativando-a ou inibindo-a. Enzimas alostérica comportam-se portanto como enzimas regulatórias em uma rota metabólica, e cuja atividade pode ser modulada em função de retroinibição ou inibição por feedback, bem como por ativação pelo precursor.
A função que define uma enzima alostérica em função do teor de seu substrato dada abaixo:
Nessa equação, nH representa o coeficiente de cooperatividade ou constante de Hill para a ligação com moléculas de S (de maneira similar à ligação de
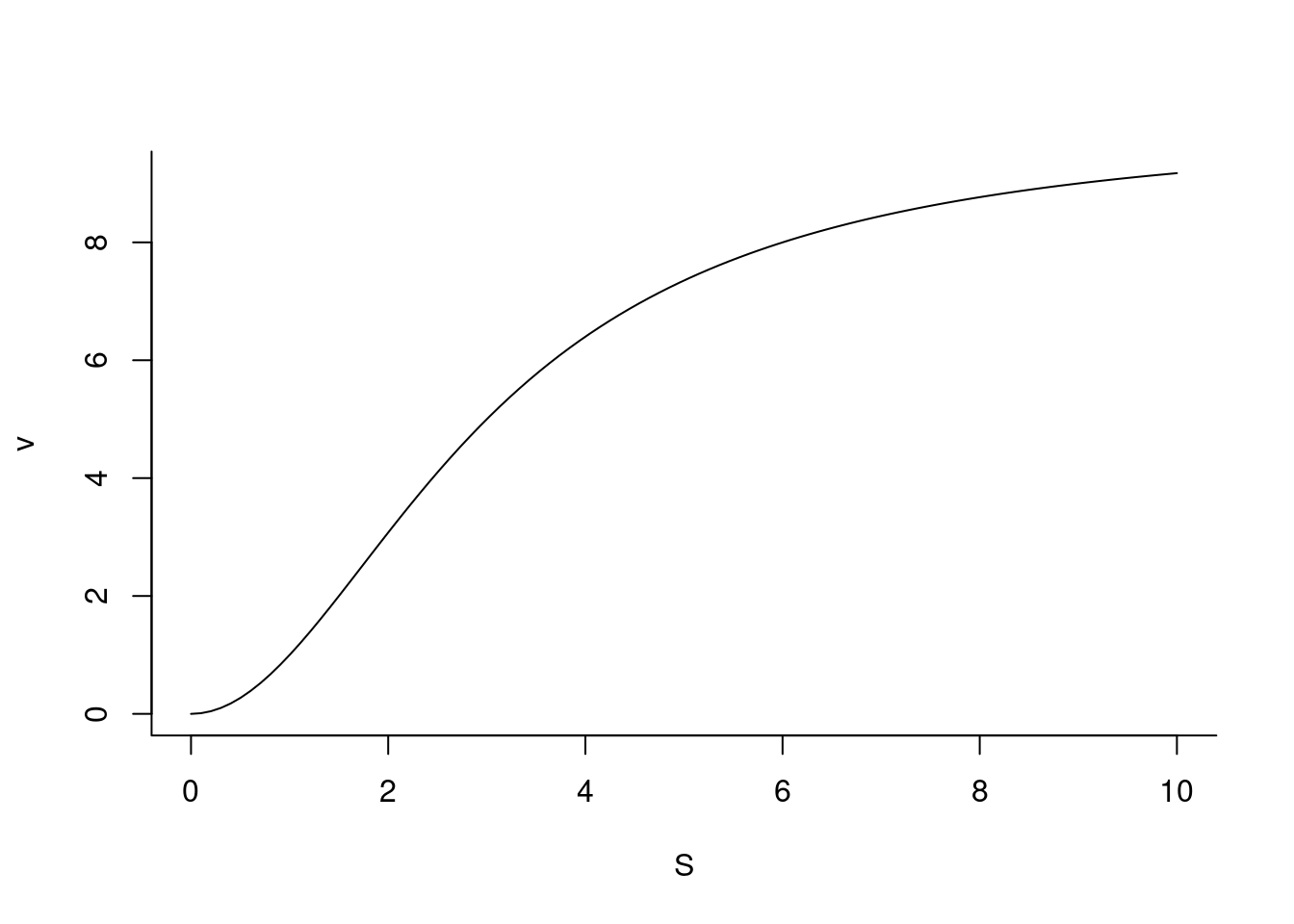
De modo geral, o valor de nH pode ser inferior à unidade (cooperatividade negativa) ou superior a essa (cooperatividade positiva).